
(Rossella Annoni & Ilaria Cavenati)
I Chirotteri rappresentano un ordine di Mammiferi tra i più diversificati e geograficamente diffusi, secondo, per ricchezza di specie, solo all’ordine dei Roditori. Le oltre 930 specie conosciute sono divise in due sottordini: MegaChirotteri e Microchirotteri.
I MegaChirotteri sono rappresentati dall’unica famiglia dei Pteropodidiae con 42 generi e circa 173 specie diverse, diffuse nelle regioni tropicali e subtropicali dell’Africa, dell’Asia meridionale, nelle isole del Pacifico fino all’Australia e alla Nuova Zelanda. Sono caratterizzati da una dieta vegetariana.
Il sottordine dei Microchirotteri è diffuso in tutti i continenti e risulta ecologicamente più differenziato, con 16 famiglie, 135 generi e 759 specie. La maggior parte dei Microchirotteri è insettivora ma vi sono specie carnivore che si alimentano di piccoli vertebrati come uccelli, rane, lucertole e anche altri Chirotteri (Megadermatidae, Nycteridae, Phyllostomidae) o di piccoli pesci e invertebrati acquatici (pochi Noctilionidae e Vespertilionidae). Esistono anche specie nettarivore e frugivore (Phyllostomidae) e persino specie ematofaghe (pochi Phyllostomidae confinati nelle regioni tropicali del Nuovo Mondo). (Fornasari et al., 1997).
I Chirotteri sono gli unici Mammiferi in grado di effettuare un volo attivo grazie alla presenza di un’ampia membrana alare e a robusti muscoli pettorali che ne permettono il movimento. Per poter rendere possibile il volo l’arto anteriore si è notevolmente modificato; infatti, tranne il pollice, tutte le altre quattro dita sono estremamente allungate e fungono da intelaiatura alla membrana alare cutanea, denominata patagio, che unisce gli arti anteriori ai posteriori e questi alla coda. Il patagio è costituito da un doppio strato di pelle all’interno del quale si trovano le ossa, i muscoli, i vasi sanguigni ed i nervi: se subisce danni di lieve entità si può rigenerare in breve tempo (una lacerazione di 10 mm può risanarsi in un mese). Nei Chirotteri il volo comporta uno sforzo muscolare notevole con un conseguente incremento della temperatura corporea; il patagio, data la sua estesa superficie di contatto con l’aria e l’attivissima circolazione sanguigna interna, funziona come un vero “radiatore”, mantenendo così entro limiti tollerabili l’ipertermia fisiologica da volo (Lanza, 1959). La forma delle ali e la velocità di volo sono estremamente varie tra i Chirotteri e sono correlate con la dieta e le abitudini di foraggiamento delle specie. Ali lunghe e strette sono tipiche di Chirotteri veloci che cacciano in spazi aperti, mentre ali più larghe o corte garantiscono una maggiore manovrabilità per le specie che cacciano in ambienti ricchi di ostacoli (Fenton, 1992).
Tutti i Chirotteri, ad eccezione dell’esotico Cheiromeles torquatus che è quasi completamente nudo, hanno la maggior parte del corpo rivestita da una soffice pelliccia più o meno fitta. Peli particolarmente lunghi e piuttosto rigidi, a funzione sensoriale (vibrisse), sono presenti in particolari aree della faccia ed altri più corti in corrispondenza dell’orecchio e del patagio.
Le mammelle sono in genere due, situate non lontano dal cavo ascellare; presenti in ambedue i sessi, appaiono ben visibili solo nelle femmine adulte prossime al parto, che stanno allattando o che da poco hanno terminato l’allattamento. Nei Rhinolophidae e in altre famiglie non europee esistono anche due capezzoli pubici che verrebbero usati dai piccoli come organi di fissazione alla madre.
Grazie a una rotazione di 180° della gamba a livello del ginocchio, il piede è rivolto posteriormente e non in avanti come in tutti gli altri mammiferi. Questo adattamento permette ai Chirotteri di appendersi ai muri.
Il muso e le orecchie sono tra le caratteristiche più sorprendenti dei Chirotteri. Molti taxa hanno ornamentazioni carnose associate al naso e alla bocca. Esse variano in forma e dimensioni e possono ricordare strutture come foglie (Phyllostomidae, Megadermatidae) e ferri di cavallo (Rhinolophidae) (Nowak, 1994). I Microchirotteri sono anche dotati di un trago e, a volte, di un antitrago (proiezioni carnose sul bordo anteriore del padiglione), assenti tra i MegaChirotteri (Vaughan, 1986).
La dentatura dei Chirotteri è difiodonte (la dentizione è formata da due serie dentarie, una decidua e una permanente) ed eterodonte (con denti diversi tra loro); quella definitiva presenta caratteristiche diverse nei vari gruppi, talora nell’ambito della stessa famiglia, soprattutto in relazione al tipo d’alimentazione. Il numero dei denti varia, nelle specie europee, dai 32 (per il genere Tadarida) ai 38 (per il genere Myotis).
L’apparato uditivo è estremamente sensibile: il padiglione auricolare può essere di forma relativamente semplice e poco sviluppato, come nei MegaChirotteri e nei Rhinolophidae, o raggiungere una notevole complicazione e dimensioni. Il genere Plecotus, infatti, presenta orecchie estremamente grandi, la cui forma e dimensione sono correlate alle abitudini predatorie. L’orecchio è fondamentale per mantenere l’equilibrio, per avvertire i suoni nel contesto sociale con altri individui, per avvertire l’avvicinarsi di eventuali predatori (Henson, 1970), nonché per l’ecolocalizzazione.
Nel 1793 l’abate Lazzaro Spallanzani scoprì che i Chirotteri, a differenza degli uccelli notturni, erano dotati della facoltà di “vedere” nel buio più assoluto e di comportarsi come se vedessero nel riconoscimento dell’ambiente circostante e delle prede. Sorpreso da questo fatto Spallanzani fece alcuni esperimenti e concluse che nei Chirotteri esiste “un novello organo o senso che non abbiam noi, e del quale in conseguenza non potremo mai avere idea”. Soltanto nel 1941 Griffin e Galambos confermarono sperimentalmente l’ipotesi di Hartridge del 1920 secondo cui i Chirotteri volando emettono ultrasuoni la cui eco, rinviata dagli ostacoli, viene captata dall’apparato uditivo; ciò permette loro di riconoscere l’ambiente circostante e le prede anche nella più completa oscurità (Figura 2.1).
Figura 2.1. I Chirotteri volando emettono ultrasuoni la cui eco, rinviata dagli ostacoli, viene captata dall’apparato uditivo (disegno di E. Comi).
Questo sofisticato sistema di orientamento viene denominato ecolocalizzazione. Tutte le specie di Microchirotteri si orientano con l’ausilio di un sistema di ecolocalizzazione mentre, tra i MegaChirotteri, solo i membri del genere Rousettus sfruttano schiocchi della lingua per ecolocalizzare le prede (Fenton, 1992). Nei Vespertilionidae europei gli ultrasuoni vengono prodotti attraverso delle contrazioni della laringe e sono emessi o con la bocca aperta o attraverso le narici. Nelle specie della famiglia dei Rhinolophidae, l’apparato nasale assume una struttura morfologicamente complessa che ha la funzione di “cono direzionale” o di “amplificatore” per una migliore concentrazione del fascio d’onda degli ultrasuoni. Gli ultrasuoni sono caratterizzati da una frequenza, da un ritmo, da una durata e da un’intensità. La frequenza corrisponde al numero di vibrazioni che si propagano nell’aria in un secondo. Le orecchie di un uomo possono percepire fino a 18.000 vibrazioni per secondo, oltre questa soglia cominciano gli ultrasuoni. I Chirotteri utilizzano le emissioni ultrasonore fino a 120.000 Hz. Possono venire emesse a frequenza costante oppure in modulazione di frequenza, cioè lungo una banda di frequenza più o meno ampia, con una discesa da una frequenza iniziale più alta a una finale più bassa: segnali con differenti caratteristiche offrono informazioni diverse sull’ambiente circostante. I Rhinolophidae emettono, per esempio, dei segnali a frequenza costante, mentre i Vespertilionidae li producono con una frequenza modulata. Gli impulsi biosonar variano durante le diverse fasi della caccia: la fase di ricerca della preda è caratterizzata da una maggiore regolarità delle emissioni; nella fase di avvicinamento, dopo che la preda è stata individuata, gli impulsi diventano più brevi e vengono ripetuti con rapidità sempre maggiore fino ad arrivare ad un ritmo molto elevato, in una brusca accelerazione finale, a breve distanza dalla preda per migliorare il dettaglio su di essa. La fase della cattura corrisponde a un breve momento di silenzio. I Chirotteri percepiscono a quale distanza si trovano le loro prede grazie all’intervallo di tempo che separa l’emissione e la ricezione dell’eco del suono emesso. La direzione è compresa grazie alla differenza di tempo che intercorre tra la percezione dell’eco dall’orecchio destro e da quello sinistro.
In tutte le specie europee il periodo degli accoppiamenti coincide con la tarda estate e l’autunno e in molti casi si protrae nei quartieri poi occupati per il letargo. Questi sono spesso gli unici periodi in cui maschi e femmine condividono i rifugi; infatti, i parti avvengono in colonie riproduttive, chiamate nursery, costituite esclusivamente o quasi esclusivamente da femmine. Un maschio può accoppiarsi con più femmine e, come nel caso di Nyctalus sp., Pipistrellus pipistrellus, Pipistrellus nathusii e alcuni Myotis, giunge a stabilire un harem di anche 10 femmine (Schober & Grimmberger, 1997). I Chirotteri, raffrontati agli altri mammiferi di piccola taglia, hanno uno sviluppo embrionale molto lento che dura dai tre ai sei mesi. Per questi animali la gestazione non ha un tempo prestabilito, ma dipende dalle risorse alimentari o dalle condizioni climatiche; per poter arrivare al sincronismo tra nascita e maggior disponibilità alimentare, i Chirotteri ricorrono a diversi sistemi. Uno di questi adattamenti è la fecondazione dell’ovulo che non si attua immediatamente dopo l’accoppiamento. Gli spermatozoi vengono conservati durante tutto il letargo nelle vie genitali della femmina restando in questo modo attivi. La maturazione dell’ovulo, la sua fecondazione e lo sviluppo embrionale si verifica solo in primavera, dopo il risveglio, quando gli insetti sono più abbondanti. Questa ovulazione differita nella zona temperata è praticata esclusivamente dai Chirotteri. Un ulteriore sistema di riproduzione utilizzata è l’annidamento differito dell’embrione. L’embrione comincia a svilupparsi durante un breve periodo ma, dopo poco, il processo si ferma: l’embrione attende le condizioni favorevoli prima di riprendere il suo sviluppo. Questo adattamento è tipico dei MegaChirotteri africani e tra i Microchirotteri europei solamente del Miniopterus schreibersii. Se si raffrontano i Chirotteri con altri piccoli mammiferi insettivori, si nota come i primi abbiano un tasso riproduttivo molto basso. La maggior parte delle femmine partorisce un piccolo all’anno, i parti gemellari sono la regola in poche specie (Pipistrellus pipistrellus, Hypsugo savii, Vespertilio murinus). I piccoli incominciano a volare a tre-quattro settimane (Pipistrellus kuhlii e Pipistrellus nathusii) o a cinque settimane per le specie più grosse (Myotis myotis e Nyctalus noctula). Lo svezzamento dura circa una decina di giorni, nei casi in cui si prolunga maggiormente, i piccoli sono indipendenti all’età di sei-otto settimane. La maturità sessuale è raggiunta all’età di un anno, con diverse eccezioni tra le femmine di molte specie, che possono essere sessualmente mature già nel primo autunno, a tre mesi di vita (Rhinolophus hipposideros, Myotis daubentonii, M. mystacinus, M. emarginatus, M. myotis, Eptesicus serotinus, Pipistrellus pipistrellus, P. nathusii, P. kuhlii e Miniopterus schreibersii). I giovani sono molto sensibili al cattivo tempo perché non hanno una riserva di grasso e mancano di esperienza. La mortalità dei piccoli dopo lo svezzamento è quindi molto alta, pari a circa il 50%. In generale si stima che solo il 30-40% raggiunga il secondo anno di vita. Alla nascita la sex-ratio è equilibrata. La mortalità dei maschi sembra essere un po’ più alta di quella delle femmine, ma questo non influenza in modo negativo la composizione della popolazione grazie alla poliginia (formazione di harem) (Schober & Grimmberger, 1991; Fornasari et al., 1997).
La particolare biologia riproduttiva dei Chirotteri rende le specie di tale gruppo particolarmente sensibili: il basso tasso riproduttivo, i tempi lunghi di gestazione, la tendenza delle femmine a riunirsi in grosse concentrazioni per il parto e per l’allevamento dei piccoli, spiegano come singoli eventi distruttivi, anche limitati ad una sola colonia, possano avere effetti sul popolamento di una intera regione (Fornasari et al., 1997).
Durante l’autunno i Chirotteri immagazzinano il grasso che gli servirà come riserva energetica per sopravvivere durante il letargo invernale. Nei Chirotteri il grasso viene accumulato principalmente intorno alle scapole, sul collo e sui fianchi. Il peso aumenta di circa 20-30% in confronto al peso primaverile. Le diverse specie durante il letargo adottano una posizione ben precisa del proprio corpo: i Rhinolophidae avvolgono il corpo con le ali come un mantello; i Plecotus piegano indietro gli enormi padiglioni uditivi molto vascolarizzati e li dispongono sotto le ali; tutti gli altri Vespertilionidae spingono le ali e l’uropatagio contro il corpo in modo da diminuire la superficie esposta all’aria. Durante questo periodo d’ibernazione si verifica una diminuzione di tutte le attività vitali, cioè delle frequenze respiratoria e cardiaca, della reattività del sistema nervoso, dell’attività muscolare, nonché delle attività endocrine e metaboliche in senso lato, con il conseguente abbassamento della temperatura corporea che viene comunque mantenuta sempre uno o due gradi sopra quella ambientale. Con tale cambiamento fisiologico delle condizioni corporee, l’animale può vivere in stato d’inattività per interi mesi superando la stagione fredda, consumando esclusivamente le riserve energetiche accumulate nei periodi di vita attiva. Per portare un esempio pratico Myotis myotis raggiunge, quando eccitato, gli 880 battiti cardiaci al minuto, a riposo si attesta intorno ai 350 battiti e, in letargo, varia tra i 18 e gli 80 battiti al minuto. La frequenza respiratoria è di circa cinque respiri al secondo da sveglio, ma, in letargo, viene ridotta a tal punto che possono esserci pause di 90 minuti tra due respiri successivi (Schober & Grimmberger, 1997). La normale temperatura corporea di un individuo in attività e di circa 40°C, i Chirotteri in ibernazione mantengono una temperatura corporea compresa tra 0° e 10°C. La scelta del rifugio per il letargo è importante: il luogo deve essere al sicuro da eventuali predatori, privo di correnti d’aria, con temperature non eccessivamente basse (comprese tra 3° e 9°C) e soprattutto con un buon livello d’umidità per impedire il disseccamento del patagio. Il letargo invernale può subire delle interruzioni per stimoli esterni, se disturbati gli animali possono spostarsi da una parte all’altra di uno stesso rifugio o anche cambiarlo, o per stimoli interni, come il raffreddamento eccessivo (sotto gli 0° C) o la vescica urinaria piena. Quando un individuo si risveglia i ritmi cardiaci e respiratori aumentano, l’apporto di ossigeno nei tessuti aumenta e la produzione di calore riprende lentamente. Solo dopo 30 o 70 minuti dal risveglio, gli animali ritrovano la temperatura normale. La durata del risveglio è maggiore se la temperatura d’ibernazione è bassa o se le riserve energetiche sono minime. In una prima fase, la produzione di calore è assicurata principalmente attraverso la combustione del grasso e in un secondo momento attraverso le contrazioni muscolari. Il tempo di letargo varia da specie a specie: va da un minimo di due giorni a un massimo di circa due mesi. Il letargo termina circa nei mesi di marzo o di aprile (Schober & Grimmberger, 1991; Fornasari et al., 1997).
Fenomeni di bioaccumulo da composti organoclorurati, metalli pesanti e policlorobifenili dovuti all’impiego di pesticidi e di altri trattamenti chimici in agricoltura e per i trattamenti del legno diventano spesso letali nel momento del risveglio dall’ibernazione. La maggior parte di tali composti viene accumulato nei tessuti adiposi. Durante il letargo e al momento del risveglio, le riserve di grasso vengono consumate rapidamente e le sostanze tossiche vengono mobilizzate in gran quantità, risultando spesso letali.
I Chirotteri effettuano degli spostamenti più o meno lunghi tra i rifugi invernali e quelli estivi. Infatti, raramente i due rifugi sono nel medesimo luogo; spesso si muovono anche solo di pochi chilometri ma cambiano ambiente: si tratta quindi di movimenti di tipo ecologico e non geografico. Le specie considerate sedentarie percorrono solitamente distanze che variano tra i 20 e 50 km tra i quartieri estivi e quelli di svernamento. La maggior parte delle specie, tra cui molte del genere Myotis, effettua migrazioni a breve distanza (da 50 a 300 km), che non necessariamente hanno un orientamento geografico preferenziale. Alcune popolazioni di Nyctalus noctula, Nyctalus leisleri, Pipistrellus pipistrellus e Pipistrellus nathusii intraprendono voli autunnali in direzione sud-ovest dai paesi nordici dove trascorrono l’estate per svernare in Francia, Svizzera e nelle aree mediterranee (spesso percorrendo anche più di 2000 Km). Nell’area di ripartizione di una specie si possono avere delle differenze di comportamento a seconda delle popolazioni coinvolte: quelle che vivono più a nord viaggiano di più di quelle che si trovano a sud. I record di distanza per i Chirotteri migratori sono detenuti da Pipistrellus nathusii e Nyctalus lasiopterus che superano spesso i 1600 km. In specie che effettuano migrazioni su lunga distanza sono state riscontrate longevità inferiori mentre, in generale, i Chirotteri sono animali considerati molto longevi: molte specie, anche di piccole dimensioni, possono superare i 20 anni di vita. I casi di longevità maggiore per le specie europee riguardano Rhinolophus ferrumequinum e Plecotus auritus, con individui sopravvissuti ben 30 anni (Schober e Grimmberger, 1991; Fornasari et al., 1997).
I Chirotteri, essendo animali gregari per la maggior parte dell’anno, utilizzano rifugi comuni, in cui gli individui appaiono aggregati. Aree ricche di habitat diversi, in cui compaiono tutte le tipologie di rifugio, presentano le maggiori concentrazioni di Chirotteri, poiché offrono molteplici opportunità di ricovero e d’alimentazione. Nel corso dell’inverno i siti di ricovero divengono luoghi dove si concentrano popolazioni d’importanza regionale o sub-regionale, poiché gli individui di molte specie tendono ad utilizzare pochi rifugi di grandi dimensioni. La maggior parte dei Chirotteri europei, in base alle diverse fasi del ciclo biologico utilizza, nel corso dell’anno, diversi tipi di rifugi che devono essere indisturbati e protetti. I rifugi utilizzati si trovano all’interno di grotte ricche d’anfratti, in fessure di muri e rocce, all’interno di edifici e in grossi alberi cavi. Al loro interno possono ospitare le nursery, cioè l’insieme di femmine gravide che si riuniscono in colonie composte da 10 a più di 100 individui, dove partoriscono ed allevano insieme i piccoli. I maschi che hanno raggiunto la maturità sessuale durante l’estate, invece, vivono spesso da soli o in piccoli gruppi all’interno di dormitori separati, sfruttati anche per l’accoppiamento. I maschi e le femmine che non hanno ancora raggiunto la maturità sessuale si riuniscono in gruppi cambiando spesso rifugio all’interno della stessa area (Schober & Grimmberger, 1991; Fornasari et al., 1997). I Chirotteri europei, in base alla tipologia di rifugio scelta, possono essere distinti in specie troglofile, specie antropofile e specie fitofile; le preferenze individuali sono da mettere in relazione anche alle caratteristiche climatiche delle diverse località e alla stagione (Schober & Grimmberger, 1991; Fornasari et al., 1997).
I Chirotteri troglofili (generi Rhinolophus, Miniopterus, Plecotus, Tadarida e buona parte delle specie del genere Myotis) sono più o meno strettamente legate ad ambienti cavernicoli come grotte e miniere ed a fessure di pareti rocciose e scogliere. Queste specie trovano in questi rifugi particolari condizioni microclimatiche come temperature basse e costanti e gradi elevati di umidità dell’aria (dal 90 al 100%). Quando individui di diverse specie occupano uno stesso grande rifugio, si nota facilmente che ognuna delle specie occupa una particolare zona della cavità, in relazione alle diverse esigenze ecologiche (Figura 2.2). In generale nelle grotte con notevole sviluppo, verso il fondo l’aria è più calda e la temperatura rimane più costante. La maggior parte delle specie si trova ad una distanza che varia dai 20 ai 60 m dall’entrata. I Rhinolophidae, che preferiscono temperature più elevate, occupano il fondo della grotta mentre il Serotino di Nilsson (Eptesicus nilssonii) e il Barbastello (Barbastella barbastellus) si trovano solitamente all’entrata delle cavità (Schober & Grimmberger, 1991; Fornasari et al., 1997). Le specie troglofile sono molto sensibili al posizionamento di strutture di chiusura (per motivi di sicurezza) nei pressi degli ingressi di grotte, miniere e gallerie. La presenza di porte o sbarre non solo può impedire agli individui l’ingresso e l’uscita dal roost, ma anche alterare il microclima (temperatura ed umidità) all’interno del rifugio, rendendolo inadatto alla presenza delle diverse specie.
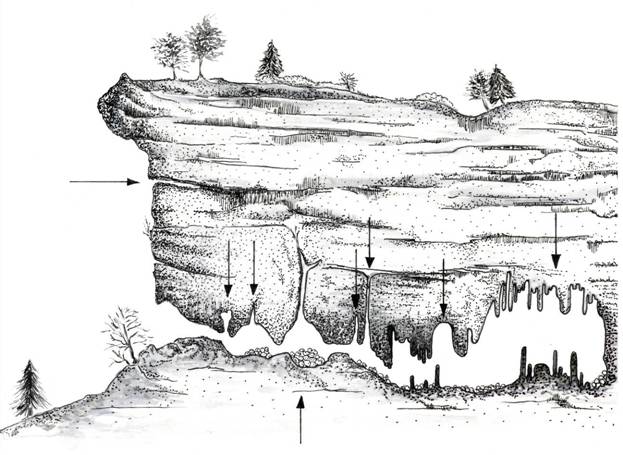
Figura 2.2. Potenziali siti di rifugio utilizzabili dai Chirotteri nelle cavità ipogee (disegno di E. Comi).
I Chirotteri antropofili, come il Pipistrello albolimbato (Pipistrellus kuhlii), il Pipistrello nano (Pipistrellus pipistrellus), il Serotino comune (Eptesicus serotinus) e parzialmente il Pipistrello di Savi (Hypsugo savii), si sono adattati a utilizzare le possibilità offerte dalle costruzioni umane (Figura 2.3). Alcune specie, come il Vespertilio maggiore (Myotis myots), utilizzano le ampie soffitte con tetti d’ardesia, materiale che si scalda molto bene immagazzinando calore; non vengono invece occupati luoghi con correnti d’aria. Anche i sottotetti delle chiese si adattano molto bene come rifugi in quanto sono luoghi senza particolari disturbi. Molte specie, grazie alle dimensioni ridotte (genere Pipistrellus), riescono ad insinuarsi in fessure larghe 1,5 cm in muri molto spessi, costruiti in mattoni o pietre combacianti e cavi all’interno (Schober & Grimmberger, 1991; Fornasari et al., 1997).
L’estrema vicinanza di colonie all’uomo spesso coincide con azioni di disturbo che, non di rado, assumono la forma di veri atti vandalici; inoltre, la necessità di effettuare ristrutturazioni spesso porta all’allontanamento di intere colonie o, in alcuni casi, all’avvelenamento degli individui a causa dei materiali utilizzati nell’intervento.
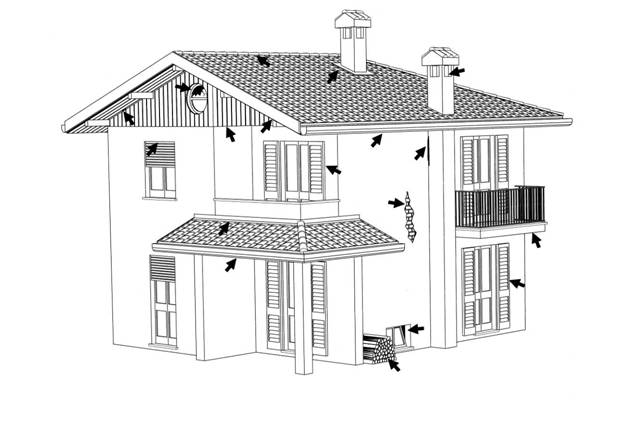
Figura 2.3. Potenziali siti di rifugio utilizzabili dai Chirotteri negli edifici (disegno di F. Farina).
I Chirotteri fitofili, quali il Pipistrello di Nathusius (Pipistrellus nathusii) ed i generi Vespertilio, e Nyctalus, occupano le spaccature naturali degli alberi o quelle prodotte da un fulmine o dallo scavo di un picchio (Figura 2.4). All’interno di queste fessure i Chirotteri si appendono sempre nel punto più alto in cui le condizioni sono più favorevoli. Qui si crea, infatti, una cupola con ristagno d’aria calda. Le nursery dei Chirotteri fitofili sono abbastanza piccole a causa del limitato spazio disponibile. I rifugi abitati dal genere Nyctalus sono facilmente identificabili poiché presentano una lunga e ben visibile striscia d’urina sotto l’ingresso in seguito all’abitudine di questi animali di svuotare la vescica prima di entrare nel rifugio (Stebbings, 1986; Schober e Grimmberger, 1991; Fornasari et al., 1997). Lembi di corteccia distaccata dal fusto della pianta possono formare fessure che offrono riparo a diverse specie di Chirotteri; la corteccia sollevata si trova in genere in alberi in fase di senescenza avanzata oppure su alberi morti o marcescenti. Generalmente non hanno dimensioni tali da offrire rifugio alle colonie riproduttive, ma vengono utilizzati in qualità di roost diurni. Utilizzano tali ripari diverse specie appartenenti al genere Myotis, quali Myotis nattereri e Myotis mystacinus. Le fessure nei tronchi d'albero forniscono rifugi analoghi a quelli offerti dal distacco di lembi di corteccia. Tali spaccature si possono produrre alla morte dell’albero stesso, o per effetto di agenti atmosferici come vento e fulmini, o per le lacerazioni prodotte dai parassiti fitofagi. Le cavità all'interno dei tronchi sono una caratteristica degli alberi vetusti; comunicano con l’esterno tramite aperture di dimensioni contenute e si possono trovare sia in alberi morti che in quelli ancora in vita, così come in alberi ancorati al suolo o sradicati. Tali cavità rappresentano per i Chirotteri i ripari più importanti, in quanto forniscono rifugi adeguati ad ospitare le colonie riproduttive di Chirotteri forestali. I rifugi costituiti dai nidi di picchio sono forse i più comuni. Sono cavità scavate all'interno del tronco dell'albero, con vani ampi almeno 10 cm e foro di ingresso di cinque-sei cm, collocati ad altezza da terra variabile tra uno e 20 m. Per tali caratteristiche un bosco maturo, ricco di alberi vetusti o morti, meglio si adatta alle necessità delle varie popolazioni di Chirotteri, grazie alla maggiore offerta di roost (Fornasari et al., 1997).
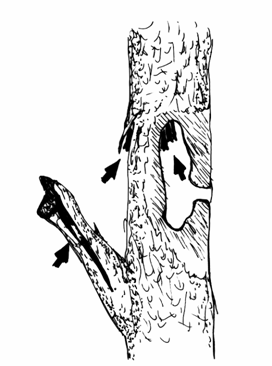
Figura 2.4. Possibili rifugi utilizzati dai Chirotteri negli alberi. I buchi scavati dai Picchi e le cavità interne vengono utilizzati fino a quando il materiale (resti di insetti, feci e pezzi di legno) accumulatosi sul fondo della cavità stessa non diviene troppo abbondante (disegno di F. Farina).
La distruzione e l’alterazione degli ambienti boschivi, corrispondenti spesso con la riduzione dei boschi più maturi e ricchi di rifugi per la chirotterofauna costituiscono una minaccia molto grave per le numerose specie che, in almeno una parte dell’anno, utilizzano questo ambiente.
I Microchirotteri utilizzano una grande varietà di habitat, non solo nella scelta dei rifugi, ma anche per quel che riguarda le zone di foraggiamento. Nelle zone temperate aree di importanza primaria sono costituite da foreste e habitat acquatici (fiumi, laghi, canali) in quanto zone naturalmente ricche di entomofauna. Soprattutto in aree fortemente antropizzate, diverse specie si sono ben adattate agli ambienti urbani, facendone habitat elettivi di caccia. Recentemente numerose ricerche hanno evidenziato l’importanza per i Chirotteri degli elementi lineari del paesaggio, quali ad esempio, margini boschivi, filari e canali, che forniscono collegamenti naturali tra le zone di rifugio e quelle di alimentazione.
È evidente che qualsiasi tipo di alterazione degli habitat naturali ha effetti negativi sulla distribuzione e abbondanza dei Chirotteri: la distruzione o alterazione degli habitat boschivi ha come conseguenza la riduzione dell’entomofauna; l’alterazione e distruzione degli habitat acquatici riduce drasticamente la disponibilità di aree di caccia per quelle specie che si alimentano direttamente sull’acqua o per quelle che utilizzano gli ambienti ripariali; la “banalizzazione strutturale” delle aree agricole, passando dalle forme di conduzione di tipo tradizionale (ecomosaici con piccoli appezzamenti coltivati, inframezzati a siepi e relitti forestali) all’agricoltura meccanizzata e intensiva (estese aree uniformemente coltivate) porta alla riduzione dell’entomofauna da un lato e alla perdita di elementi “lineari” dall’altro.